
The two-headed bacterium

I like to see categories as fish nets we use to capture ideas. We classify things into categories like individuals, nation or species, and of course it is all arbitrary and doesn't correspond to anything in the real world. But categories still form useful chunks we can use to make sense of the world. Furthermore, here is a fun exercise: introduce arbitrary changes in the categories, and see what the world looks like through this new lens. As I will argue, there are plenty of things to be discovered this way. Use the standard fish nets, and you get a standard understanding of the world. Try to use slightly larger or smaller nets, and maybe you will discover things you had never noticed before.
Take the individual, for example. One bacterial cell contains exactly one genome and all the necessary equipment to replicate it. Using our human-derived intuition of what makes an individual, it makes sense to see bacteria as unicellular organisms, meaning that one cell = one individual. If you visit the wiki page on prokaryotes (the larger group that encompasses bacteria and archaea) the first thing you hear is that they are unicellular, as if it were the most important thing about them. However, bacteria are so weird, so different from us, that it makes little sense to describe them using the categories we invented while observing humans.
Let's explore the strange and surprising processes that are uncovered when you change your definition of the individual to make it either wider, or narrower. First, I will start with a hot take: each bacterial lineage is one big multicellular individual. Then I will move on to the super-hot-magma-take: each bacterial cell is actually made of two distinct individuals fused together, facing in opposite directions.
Bacteria as multicellular organisms
First, let's make our definition of the individual arbitrarily broader, and consider that the whole bacterial culture, descending from a single ancestral cell, is one individual. Is there anything interesting to see here? For starters, some behaviors of bacterial cells don't really make sense as individuals. For example, bacterial cells regularly perform what could only be described as bacterial sacrifice.
The Kelly criterion in prokaryotes
Content warning: bacterial sacrifice
Antibiotics were already in the environment long before humans started using them, usually secreted by other micro-organisms who want to take your precious nutrients for themselves. Imagine being a bacterium growing peacefully – there is always a risk that some bastard fungus will put their filthy pterulone, sparassol or strobilurin in your soup. Fortunately, bacteria figured out a solution: enemies can't stop you from growing if you are already not growing.
In its simplest form, this works because the antimicrobial compound needs to be actively incorporated in the growth machinery to cause trouble. Think of a grain of sand being caught in a clockwork mechanism and breaking everything – if the mechanism is stopped, the grain of sand doesn't enter, and you can resume operation later once the grain of sand has been blown away. Obviously, the drawback is that the bacterium is no longer growing, which kind of defeats the whole point. This is why bacteria have invented what we humans know as the Kelly betting system.
Say a gambler bets on something with 2:1 odds, so if she wins the bet, she gains twice as much as what she invested. She know she has a 60% chance of winning, so the most profitable strategy is of course to invest 100% of her money every time – this way, she maximizes the return of every winning bet! But obviously this is bad, because eventually she will lose a bet, and then have zero monies remaining. For bacteria, this is like having 100% of the cells growing as fast as they can. This maximizes the population growth rate, until the aforementioned bastard fungus secretes some pleuromutilin or whatever and then the entire population takes it up and goes extinct. To avoid this, our gambler should invest only a fraction of her money on each bet, so her funds still grow exponentially (albeit at a slower rate) but in case of loss she still has some funds to continue. For bacteria, this means always having a small fraction of the population that stops growing, as a backup. This is essentially the bacterial population betting on whether there will be antibiotics in the close future. From the perspective of an individual cells, both situations are bad – either you stop growing, while your friends quickly outnumber you by orders of magnitudes and you practically disappear, or you are part of the growing fraction and eventually you die from antibiotic overdose. But if you look at the entire colony, you can see the two sub-populations as two essential parts of a single organism, that figured out some slick decision theory techniques long before the species of John L. Kelly even evolved a brain.
Eating the corpses of your siblings
Content warning: eating the corpses of your siblings.
Similarly, one puzzling feature of bacteria is that they sometimes commit apoptosis. This happens, for example, when food is scarce – some cells may spontaneously explode so that other cells can feed on their remains, increasing the chances that at least one of them will make it out alive when resources come back. If you see each cell as an individual, that is weird, and does not fit well with anything methodological individualism would predict. But if you see the whole colony as the individual, then it is just like your good old typical apoptosis – just like, in the fetal stage, your fingers were all connected by cells until some of them honorably committed seppuku so you get born with fingers instead of webbed paws.
(One fascinating thing with bacterial apoptosis is that every cell which ever activated these pathways is dead. Thus, if you look at a currently living bacteria, at no point in billion years of evolution did this pathway ever activate in any of its ancestors. Not even by chance. The entire mechanism evolved and improved only by correlation with other cells, without ever activating in the lineages we can now see.)
Action potentials in biofilms
As a third exhibit of things bacteria do that definitely don't look like unicellular behavior, there is the recent discovery that some bacteria, after organizing themselves as a biofilm, are able to communicate with each other using electrical waves. The way it works is remotely similar to the action potentials we see in neurons. At a resting state, cells are filled with potassium ions, which makes them electrically polarized. Whenever the polarization disappears, ion channels in the envelope open up, and the potassium ions all exit the cell into the extracellular environment. This, in turns, cancels out the polarization of neighboring cells. The result is this:
Supposedly, this mechanism makes sure the outer bacteria will stop eating from time to time, so the nutrients can diffuse all the way to the center and prevent the interior cells from starving. If this does not make you scream "multicellular!", I don't know what will.
In short, rather than being just individual cells fighting against each other, bacteria have evolved hard-wired mechanisms that only make sense if you consider the dynamics of the whole colony. A microbiologist could spend her entire career building a perfect model of one bacterial cell, but she would still be far from understanding all facets of the organism. Oh, and if you are ready to hear a similar point about humans (that is, human communities are multi-body individuals), get your largest fish net and check out this review. I will continue with bacteria, because we have barely scratched the lipopolysaccharide of their weirdness.
Bacterial cells are two-faced pairs of individuals
Now, let's see what happens with a much narrower definition for an individual. Even narrower than a single cell. Put down that extra-large "big game"-rated landing net and bring the tweezers.
Here is our new definition: an individual is what happens between a birth event and a death event. Now we need to find definitions of birth and death that apply to bacteria. Let's say, a birth event is when a mother cell divides into two daughters (specifically, cytokinesis). A death event is when a cell is irreversibly broken, is torn apart or becomes too damaged to grow. We have a simple and precise definition, now we can look at bacteria and pick apart the individuals.
One generation goes as follows:
The cell extends and roughly doubles in length
The middle of the cell constricts and two new poles are constructed
The cell divides and you get two cells. Each of them has one old pole that was already there in the previous generation, and one shiny new pole:

Where is the individual here? Now you understand why I came up with that bizarre birth-death definition. First, let's number the poles according to their age (in generations).

But what if bacteria age? It turns out that, yes, bacteria age. After a number of generations, old poles accumulate damage. Depending on the growth environment, they may still be fine, or grow slower, or explode in an effusion of bacteria blood. To reduce clutter, I'll consider that poles have a lifespan of 3 generations, and then the cell is dead (in real life, they hold for much longer, but that wouldn't be sketchable).
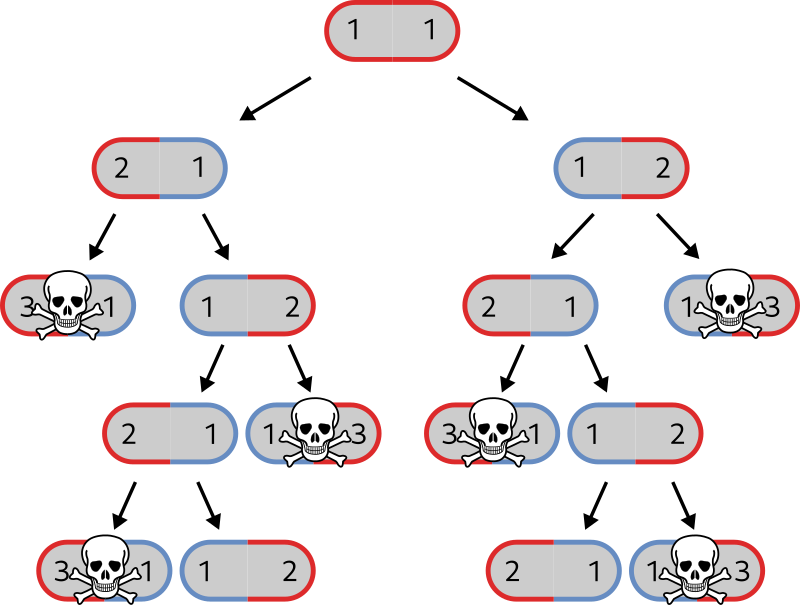
Coming back to our custom, "birth-to-death" definition of an individual, you can see that each cell is actually made of two of them – one on the left, one on the right.

Here they are very short-lived and die after three generations, but in real life these "half-bacteria" live for much longer, perhaps hundreds of generations if the conditions are not too bad. But the principle remains the same, there are just a lot more of these diagonal individuals.
Using your ancestors as trashcans
Content warning: yeah, that.
But wait, there is more. As I said, in nice conditions the poles can grow basically forever. Yet they still exhibit aging. And yes, this is all sane and coherent. This is where the titles of the papers become really spooky (Age structure landscapes emerge from the equilibrium between aging and rejuvenation in bacterial populations or Cell aging preserves cellular immortality in the presence of lethal levels of damage), showing how far we are from our typically construction of the individual.
To put it very briefly, take the sketches above where half of the cell is young and half of the cell keeps getting older. Old material accumulates in the old pole, so those cells keep growing slower and slower after each generation. Now add some mixing to it: every generation, the older pole gets a little bit of fresh material, and the younger pole gets a little bit of old material. Eventually the old pole reaches an equilibrium when the new material their inherit exactly compensates the damage from aging. As there is the same thing, reversed, for the young pole, you end up with two attractors:

What is the importance of this? There may be no importance at all, since the old cells are quickly outnumbered by young cells so they only represent a tiny fraction of the colony. However, there is also some evidence that all kinds of garbage, like misfolded proteins or aggregates, tend to accumulate in the old pole. Perhaps this ensure that at least some cells in the population will be in perfect shape, so in case of trouble, they have a good chance of having at least one survivor (a bit like North Korea preparing a team for the Math Olympiads).
But this, of course, brings us back to collective, multicellular behavior. Life is too complicated to fit in a single fish net.